LATERALIZATION OF FUNCTIONS AND SPATIAL COGNITION IN TUFTED CAPUCHIN MONKEYS (Cebus apella)
INTRODUCTION
An emerging perspective related to lateralization in nonhuman primates is based on the recognition of an apparent lack of any relationship between the use of the hands and lateralized cognitive functions (Hamilton & Vermeire, 1988b). Few studies have found an association between hemispheric learning and hand preference (Hamilton, 1983; Horester & Ettlinger, 1985). From an evolutionary point of view, the absence of specific hand preference in populations of nonhuman primates does not necessarily imply that the brains of such primates are bilaterally symmetric for all functions. In fact, hemisphere-specific functions, as seen in humans, can operate independently from the use of the hands. Yet the specific relation between lateralized functions and hand use in nonhuman primate species can differ from the function found in humans.
Considering these possibilities, researchers have begun to focus their studies on lateralization functions, in which there are great cognitive similarities between human and nonhuman primates, especially in areas such as auditory processing (Pohl, 1983), oral communication (Petersen et al., 1978), facial expression (Hamilton & Vermeire, 1988a), and spatial learning (Hamilton, 1983; Jason et al., 1984).
The data currently available suggest that there is a preference for the left hemisphere for processing certain types of visual-spatial (i.e., geometric) information, whereas the great apes, with language training, have presented a strong advantage of the right hemisphere in visual-spatial discrimination (Hopkins & Morris, 1989) and form recognition (Hopkins et al. 1990).
These findings might be related to differences among species. However, this suggestion clearly requires further investigations, using similar procedures and stimuli in different species, in order to draw any conclusions. In addition, most research on lateralization of cognitive functions has involved only a few animal species: rhesus monkeys (Macaca mulatta); chimpanzees (Pan troglodytes); and gorillas (Gorilla sp.) Therefore, it is essential to extend this investigation to other primate species.
The objective of the present study is to explore the above-mentioned issue in a species of New World monkey, the tufted capuchin (Cebus apella).
MATERIALS AND METHODS
Subjects:
Two 8-year-old (adult) male tufted capuchins, named Tiquinho (Tiny) and Alemão (German), were used. Both individuals were born in the wild and taken into captivity when they were still juveniles. They were then placed together with other individuals on islands. During the experiment, the individuals were transferred to isolated enclosures and held in cages (2 m × 2 m × 2 m).
Materials:
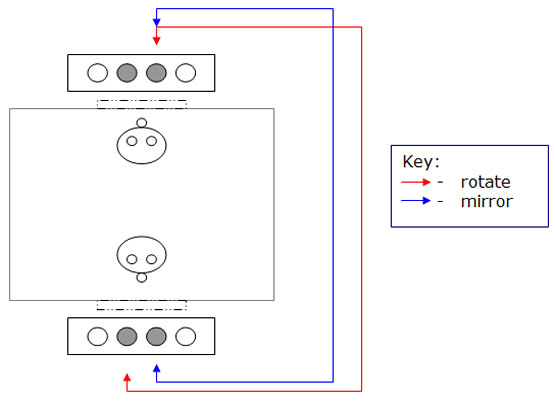
- two tables (60 cm × 25 cm × 60 cm), with wheels (to be moved) and four depressions (2 cm deep, 8 cm in diameter, 8 cm apart).
- two bowls that were identical in form and color, together with two bowls of identical form but different in color (one white and one blue).
- wooden screens that covered the cages in order to prevent the individuals from seeing out or to limit their field of vision.
- These screens were composed of articulated modules that could be opened, like windows, during the training and testing phases.
- chair for the experimenter in front of the test table.
Procedures:
One object-choice task was employed, in which the subject had to choose one of two bowls seated in two of the depressions in the center of the table in order to recover bits of food previously hidden beneath one of the bowls. In the training sessions, the subjects learned on which side the food was hidden. The task was considered learned when the subject met the criterion of 90% correct answers in a series of trials conducted in 10-min blocks. The test sessions, composed of five blocks of six trials each, with an interval of at least 90 minutes between each block, then began.
Every trial consisted of showing the table, placing it within the reach of the subject and oriented as it had been in the training. The food was hidden beneath the bowl, also positioned as in the training. Soon after, the window on the opposite side of the cage was opened, and an identical table was presented to the subject. The side chosen by the subject in the attempt to recover the food, as well as the time spent in completing this task, was noted by the experimenter. This procedure was conducted on both the right and the left sides.
The tasks were performed under two circumstances:
a-) the bowls seated in the depressions were of the same form and color.
b-) the bowls seated in the depressions had same form but were of different colors (white and blue), and the food was hidden beneath the blue bowl.
In the second circumstance, when the subject went to the opposite side from where the training had been performed, it found the blue target in a symmetric position in relation to the target on the other side (mirrored position) (Diagram 1).
RESULTS
The summary of the performance of the subjects in the first phase (phase a) is shown in Table 1.
Table 1. Result of the search task under circumstance (a): two identical bowls.
|
|
||||||
| Right Side | Left Side | |||||
|
|
|
|||||
| Subject | Rotated | Mirrored | Time(s) | Rotaded | Mirrored | Time(s) |
| (x ± SE) | (x ± SE) | |||||
|
|
||||||
| Tiny | 26 | 4 |
0.07 ± 0.02 |
12 | 18 | 1.88 ± 0.4 |
| German | 23 | 7 | 0.7 ± 0.23 | 14 | 16 | 0.15 ± 0.04 |
|
|
||||||
| SE: standard error | ||||||
The results show that, considering the fact that the bowls were identical, and the individual only had spatial information to guide the choice, the individual tended to rotate its reference system, that is, the target bowl was positioned on its right side during the training phase (Binomial test, Unidirectional: Tiny: n = 30, p < 0.001; German: n = 30, p < 0.003). However, the results observed when the target was on the left side (from the perspective of the subject) in the training phase do not indicate any particular trend (Binomial test, Bidirectional: Tiny: n = 30, p > 0.22; German: n = 30, p > 0.5).
The results of the second phase (phase b) are summarized in Table 2.
Table 2. Result of the object-choice task under circumstance (b): the bowls were of different colors (one blue and one white), and the food was hidden beneath the blue bowl, which was in a symmetric position in relation to the opposite (mirror) side.
|
|
||||||
| Blue - Right Side | Blue - Left Side | |||||
|
|
|
|||||
| Subject | Rotated | Mirrored | Time(s) | Rotaded | Mirrored | Time(s) |
| (x ± SE) | (x ± SE) | |||||
|
|
||||||
| Tiny | 27 | 3 |
2.12 ± 0.28 |
5 | 25 | 1.75 ± 0.17 |
| German | 22 | 8 | 0.56 ± 0.07 | 10 | 20 | 0.51 ± 0.09 |
|
|
||||||
| SE: standard error | ||||||
The results show that, when the target was placed on the right side of the subject in the training phase, there was a tendency toward rotating its reference system, despite the color variable. Therefore, the geometrical information was preferably used in deciding where to look for food (Binomial test, Unidirectional: Tiny: n = 30, p < 0.0001; German: n = 30, p < 0.008).However, when the target was placed on the left side of the subject, there was a tendency toward using the color as a guide for the choosing process (Binomial test, Unidirectional: Tiny: n = 30, p < 0.001; German: n = 30, p < 0.05).
DISCUSSION
The results suggest that the ability of these monkeys to rotate their spatial reference system by 180° is asymmetric in relation to the right and left sides. The monkeys tended to rotate their spatial reference system when the food was beneath the bowl placed on the right side, and they did not show any particular tendency when the food was on the left side.
The introduction of the color variable had two different effects on subject behavior: when the food was placed in a position that favored the rotation of the spatial reference system, i.e., on the right side, the color, understood as a clue to find the hidden food, was ignored, and the subject resorted to using only the geometrical information to find the food. However, when the food was in a neutral position, i.e., on the left side, the subjects tended to use the color as a guide.
These data suggest asymmetry in the representation of space and lateralization of the cognitive function in tufted capuchins. In addition, this seems to indicate that the type of clue – geometrical or environmental – that the monkeys preferably use to represent the space varies in relation to the right and left sides. Further experiments have been planned in order to analyze the possible involvement of differential hemispheric learning in this type of spatial task.
REFERENCES
Hamilton, C. R. & Vermeire, B. A. 1988b. Cognition, not handedness, is lateralized in monkeys. Behavioural and Brain Sciences, 11: 723-25.
Hamilton, C. R. 1983. Lateralization for orientation in split-brain monkeys. Behavioural Brain Research, 10: 399-403.
Hamilton, C. R., & Vermeire, B. A. 1988a. Complementary hesmiferic specialization in monkeys. Science, 242: 1694-96.
Hopkins, W. D. & Morris, R. D. 1989. Laterality for visual-spatial processing in two language-trained chimpanzees. Behavioral Neuroscience, 104: 227-234.
Hopkins, W. D., Washburn, D. A. & Rambaugh, D. M. 1990. Processing of form stimuli presented unilaterally in humans, chimpanzees (Pan troglodytes), and monkeys (Macaca mulatta). Behavioral Neuroscience, 104: 577-82.
Horester, W., & Ettlinger, G. 1985. Na association between hand preference and tactile discrimination performance in the rhesus monkey. Neuropsychologia, 23: 411-13.
Jason, G. W., Cowey, A., & Weiskrantz, L. 1984. Hemispheric asymmetry for a visual-spatial task in monkeys. Neuropsychologia, 22: 77-84.
Petersen, M. R., Beecher, M. D., Zoloth, S. Moody, D. B., & Stebbins, W. C. 1978. Neural lateralization of species specific vocalizations in Japanese macaques (Macaca fuscata). Science, 202: 324-27.
Pohl, P. 1983. Central auditory processing: Ear advantages for acoustic stimuli in baboons. Brain and Language, 20: 44-53.